- Home
- News
- Spotlight on Science
- Phonons reveal molecular-level...
Phonons reveal molecular-level organisation of phase-separated lipid membranes
10-03-2020
Interaction of lipid species in a cell membrane remains poorly understood despite extensive studies. Inelastic X-ray scattering has provided new evidence revealing the role of strongly-interacting transient lipid pairs in the organisation of liquid-liquid phase separation in lipid membranes.
The cellular plasma membrane is a highly-complex and heterogeneous environment for which lipids are sorted biochemically within the cell. The lipid rafts hypothesis proposes that preferential association of different lipids with sterols and proteins is responsible for a multitude of the cell membrane’s biological functions. Cholesterol, the major non-polar lipid, is thought to define the integrity of a cell membrane and to trigger the liquid-liquid phase separation into liquid-disordered (Ld) and liquid-ordered (Lo) phases. The latter is considered a fundamental property of the membrane, which governs a variety of interaction pathways of a cell with the extracellular environment. Several models were invoked to describe the lipid interactions in membranes with coexisting liquid–liquid phases, however, they fall short of drawing a consistent molecular picture of a cell membrane at the nanoscale, especially in the presence of different lipid species (saturated, unsaturated, or polyunsaturated) and proteins.
The vibrational processes in single component biological membranes have recently been studied using inelastic X-ray scattering (IXS) [1]. Highly-dynamic, nm-sized, localised lipid clustering in lipid membranes was observed to form and relax on a picosecond time scale. Such clustering, or local chain ordering, was argued to define the mechanism of transmembrane solute diffusion. Here, a unique picosecond-nanometre time–space window of IXS has been exploited to study the lipid-lipid interaction in multicomponent model membranes at the molecular level. Using high-resolution IXS measurements of binary mixtures of DPPC (1,2-dipalmitoyl-sn-glycero-3-phosphocholine) and cholesterol and ternary mixtures of DPPC, cholesterol and DOPC (1,2-dioleoyl-sn-glycero-3-phosphocholine) or POPC (1-palmitoyl-2-oleoyl-glycero-3-phosphocholine), two important discoveries were made. First, an optical phonon mode was observed experimentally for the first time in lipid mixtures. Second, the measurements revealed the opening of a phononic gap in the optical mode. The size of the gap depends on whether unsaturated lipids are present in a mixture.
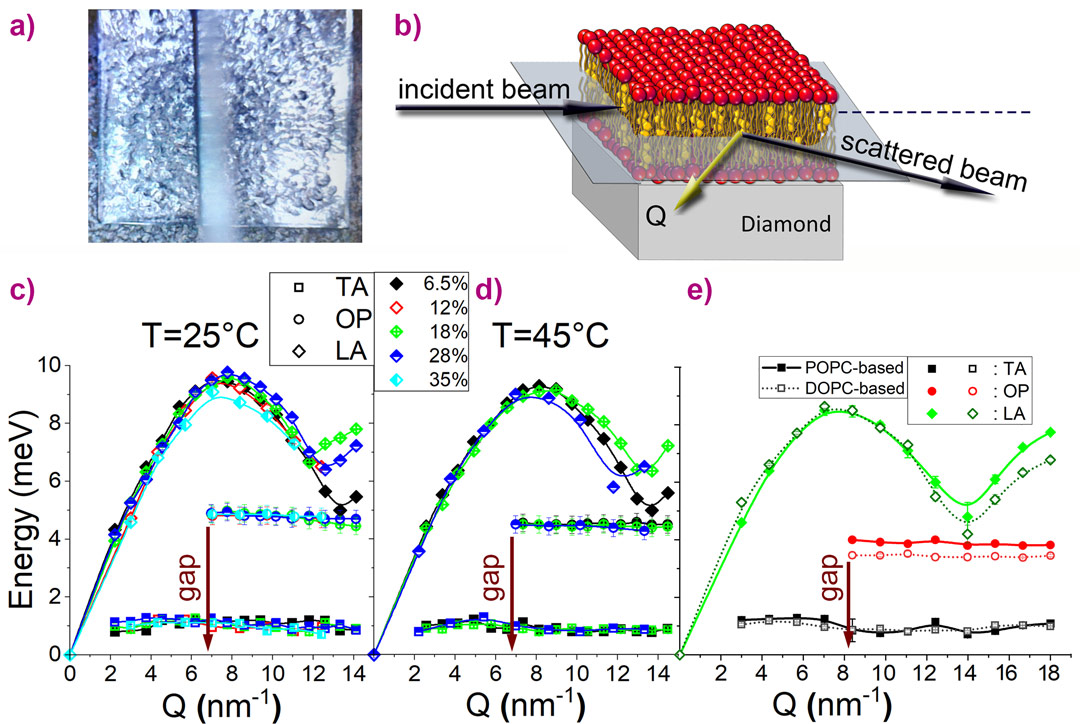
The measurements were performed at beamline ID28, collecting data for the binary mixtures at 25°C and 45°C and for the ternary mixtures at 37°C. Complementary measurements were performed at beamline BL35XU at SPring-8. Figure 1 illustrates the scattering geometry of the IXS experiment, the experimentally obtained phonon dispersion curves, and the schematic representation of the phonon propagation in the sample.
 |
|
Figure 1. (a) A real image (the top view) of a lipid multilayer deposited on top of a 4.5 × 4.5-mm2 diamond substrate. (b) The scattering geometry of the IXS experiment: the scattering vector Q remained within the plane of the lipid layers, ensuring that only the in-plane dynamics were probed. (c-e) The dispersion curves for DPPC-Chol mixtures (c-d) at different mol% of cholesterol as indicated in the legend taken at 25°C and 45°C and for the POPC(0.5):DPPC(0.3):Chol(0.2) (POPC-based) and the DOPC(0.3):DPPC(0.4):Chol(0.3) (DOPC-based) ternary systems (e) taken at 37°C. The solid lines are guides to the eye only. (f) The schematics of the phonon propagation in the sample (lipid displacements are greatly exaggerated): the longitudinal acoustic mode is represented as a compression wave propagating through a sheet of lipid molecules, and the transverse acoustic mode is represented as a shear wave with the in-plane molecule displacements. The non-dispersing optical mode located close to 5 meV is explained in Figure 2. The phononic gap in the optical mode is indicated by an arrow. |
The optical phononic (OP) mode, or standing waves, in the binary and ternary systems originates from the out-of-phase collective movements of neighbouring DOPC, POPC, DPPC, and cholesterol molecules about their centre of mass (Figure 2). The observation of the phononic gap on the OP mode is highly significant, as its appearance in materials is due to the finite size effect [2]. In lipid mixtures, a low momentum transfer phononic gap is a manifestation of the presence of strongly-coupled, nanoscaled lipid units in the lipid mixture (therefore, the finite size effect), the size of which correlates directly with the phononic gap (Figure 2).
 |
|
Figure 2. Schematic representation of the Ld-Lo phase coexistence, where up to three lipid pairs nucleate into nanoscaled lipid units, or nanodomains. All DPPC or Chol molecules within the purple region depicting a nanodomain move coherently in opposite directions indicated by red and green arrows (all DPPC move to the left and all Chol move to the right). As a result of coherent antiphase oscillations of DPPC and Chol species about their static centres of mass (black circles), the OP mode is supported over the length scale inferred from a value of the phononic gap. |
The results provide an important insight into the mechanism of the raft formation and lipid-lipid interaction driving the liquid-liquid phase separation. Specifically, the observation of the OP mode and the phononic gap suggests that the Lo phase, which is believed to be related to lipid rafts in the plasma membrane, is inhomogeneous on the nanoscale and is represented by a set of transient discrete nanoscaled units, or functional lipid pairs (up to 1 nm in diameter). Mobile in nature, lipid pairs can freely diffuse and further coalesce to form stabilised rafts of larger size through transient recruiting of other lipids or proteins. Noteworthy, unsaturated lipids proved to play an equally important role in driving the liquid-liquid phase separation, which historically was attributed to the attractive interaction between saturated lipids and cholesterol and exclusion of (poly)unsaturated lipids into the Ld phase. The observation of lipid pairs in ternary systems points to an interplay between repulsive and attractive interactions of unsaturated, saturated lipids and cholesterol in the raft formation. These results support the notion that individual lipids can allosterically regulate the function of transmembrane proteins. The ultrafast dynamics of functional lipid pairs (the energy of the optical modes) operates at the same timescale as relaxation processes of the transmembrane proteins, which highlights the lipid pairs’ role as relaxation channels within the membrane plane.
Principal publication and authors
Functional lipid pairs as building blocks of phase-separated membranes, D. Soloviov (a,b,c,d), Y.Q. Cai (e), D. Bolmatov (f,g), A. Suvorov (e), K. Zhernenkov (h,b), D. Zav’yalov (i), A. Bosak (j), H. Uchiyama (k), M. Zhernenkov (e), Proc. Natl. Acad. Sci. U.S.A. 117, 4749-4757 (2020); doi: 10.1073/pnas.1919264117.
(a) Moscow Institute of Physics and Technology, Dolgoprudny (Russia)
(b) Joint Institute for Nuclear Research, Dubna (Russia)
(c) Taras Shevchenko National University of Kyiv, Kyiv (Ukraine)
(d) Institute for Safety Problems of Nuclear Power Plants of NAS of Ukraine, Chornobyl (Ukraine)
(e) Brookhaven National Laboratory, Upton (USA)
(f) Oak Ridge National Laboratory, Oak Ridge (USA)
(g) University of Tennessee, Knoxville (USA)
(h) Forschungszentrum Jülich GmbH, Garching (Germany)
(i) Volgograd State Technical University, Volgograd (Russia)
(j) ESRF
(k) SPring-8, Hyogo (Japan)
References
[1] M. Zhernenkov, D. Bolmatov, D. Soloviov, K. Zhernenkov, B.P. Toperverg, A. Cunsolo, A. Bosak, Y.Q. Cai, Nat. Commun. 7, 11575 (2016).
[2] D. Bolmatov, M. Zhernenkov, D. Zav'yalov, Y.Q. Cai, A. Cunsolo, A. J. Phys. Chem. C 120, 19896-19903 (2016).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.